



Tyrannosauridae
Tyrant dinosaurs
Thomas R. Holtz, Jr.

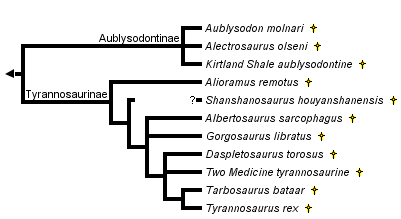

This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxPhylogeny based on Holtz (in press a, b). Please refer to the page on Tyrannosaurid Systematics for synapomorphies supporting this tree. See Taxonomy for definitions of Aublysodontinae and Tyrannosaurinae.
Introduction
Tyrannosauridae is one of the best-represented clades of large bodied theropods in the fossil record. Five species in three to five genera are known from multiple relatively complete skeletons representing different ontogenetic stages: all five species include individuals with femoral lengths of over 1000 mm. Several other tyrannosaurid taxa are known from more incomplete remains.
Tyrannosaurid tooth remains were reported by Leidy in 1856; and partial skeletal material was described in the late 19th Century by Cope (1892) and Marsh (1890, 1892). Tyrant dinosaur anatomy has been described in more detail by Lambe (1904, 1917), Osborn (1905, 1906, 1912, 1916), Parks (1933), Russell (1970), Maleev (1974), Molnar et al. (1990), Molnar (1991), and Carr (1999).
Tyrant dinosaur paleobiology has been the subject of numerous technical and popular studies and speculations, due to the fact that they are known from relatively complete fossils of large to gigantic size and highly derived anatomy.
Some authors (Lambe 1917, Colinvaux 1978, Halstead and Halsted 1981, Barsbold 1983, Horner and Lessem 1993, Horner 1994, Horner and Dobb 1997) have suggested that tyrannosaurids in general or Tyrannosaurus rex in particular were obligate scavengers, incapable of taking down live prey. However, Farlow (1993) and Holtz (in press c) have found these theoretical, ecological and ecomorphological arguments wanting at present. Instead, as is the case with most large-bodied terrestrial carnivores, tyrant dinosaurs probably used both scavenging and predation in order to obtain their food. Carpenter (2000) has documented an adult specimen of the hadrosaurid Edmontosaurus annectens with a bite mark attributable by morphology and stratigraphy to Tyrannosaurus rex: because this injury shows subsequent regrowth of bone, this wound was inflicted on a living animal rather than a carcass.
Tyrannosaurid dentition is markedly heterodont, perhaps indicating a partition of different sectors of the jaw for different function: scraping for the incisiform premaxillary teeth versus tearing and crushing for the lateral teeth, for example. Despite statements to the contrary (Feduccia 1999), the dentition of tyrannosaurids is not similar to those of other theropods. Instead, the incrassate (labiolingually expanded) lateral teeth of tyrannosaurines have greater mediolateral and craniocaudal bending strength than those of other theropods (Farlow et al. 1991, Holtz in press c). Erickson and Olson (1996), Erickson et al. (1996), Molnar (2000), Hurum and Currie (2000), and Holtz (in press c) have argued on tooth mark and biomechanical evidence that tyrannosaurids were capable of producing a greater crushing bite than other large theropods, such as allosauroids and abelisaurids. A very large coprolite from the Scollard Formation, probably referable to Tyrannosaurus rex, is nearly half macerated ornithischian bone by volume (Chin et al. 1998), consistent with the crushing bite model.
Tyrannosaurid forelimbs are extremely reduced, both in proportion to the body and in terms of digit development. In contrast, the hindlimbs of tyrant dinosaurs (and in particular the tibiae and metatarsi) are elongate compared to those of most other theropods of the same body mass. Indeed, the hindlimb proportions of tyrannosaurids, ornithomimosaurs, and the gracile ceratosaur Elaphrosaurus form a continuous allometric trend, suggesting that the tyrant dinosaurs had a greater cursorial ability than other large-bodied theropods (such as allosauroids and spinosauroids) (Holtz 1995, Carrano 1998, Paul 2000). However, Farlow et al. (1995) have calculated that at extremely high speeds an adult Tyrannosaurus rex would be subjected to fatal damage if it fell, suggesting that very large individual tyrannosaurids may not have been as cursorial as younger or smaller specimens.
Currie (2000) reports the discovery of at least nine individuals of Albertosaurus sarcophagus from the same quarry; these were of different ontogenetic stages, suggesting that at least some tyrannosaurid taxa may have been gregarious.
Characteristics

Nearly complete skeleton of a subadult Gorgosaurus libratus, from the Royal Tyrrell Museum of Palaeontology. Photograph © 1998, Thomas R. Holtz, Jr.
Tyrannosaurid anatomy is highly apomorphic (Holtz in press a, b). The following features are present in tyrannosaurids basally, and in some but not all of their potential outgroups. As such, depending on the phylogenetic position of the tyrant dinosaurs, these may be diagnostic features of Tyrannosauridae or synapomorphies with that taxon's closest relatives (Holtz 1994, 2000, in press a, b, Sereno 1999):
- Incisiform premaxillary teeth
- Nasals narrow caudally behind external nares
- Lacrimal-frontal contact present; prefrontals reduced
- Squamosal recess
- Quadrate height 40-50% of skull height at mid-orbit
- Coracoid dorsoventral length 5 times or more coracoid glenoid diameter
- Radial distal end flattened craniocaudally
- Distal carpals of adults poorly formed and lack trochlear surfaces
- Pollex ungual subequal to ungual of digit II in size
- Preacetabular notch on ilium
- Distal end of iliac pubic peduncle oriented horizontally
- Iliac supracetabular crest transverse width straight
- Dorsal surfaces of iliac blades very close to the midline
- Pronounced semicircular scar on the caudolateral surface of the ischium, just distal to the iliac process
- Ischial foot absent
- Femoral anterior trochanter as tall as greater trochanter
- Anterior and greater trochanter junction at distal end of anterior trochanter
- Tibiae and metatarsi elongate relative to other theropods of same femoral length (Holtz 1995)
- Calcaneum distal articular width 10% of maximum transverse width of astragalus
- Presence of an arctometatarsus (proximal end of metatarsal III reduced to a solid splint) (Holtz 1995)
- Metatarsals II and IV contact at mid-shaft on plantar surface
Additionally, the following features are diagnostic of Tyrannosauridae:
- Ventral ramus of premaxilla taller dorsoventrally than long rostrocaudally
- Nasals fused
- Marked ventral curvature of maxilla
- Squamosal-quadratojugal flange intruding rostrally into infratemporal fenestra, nearly contacting rostral margin
- Well-developed sagittal crest on dorsal surface of parietals
- Supratemporal fossa occupies at least the laterocaudal third of frontal
- Caudal surangular foramen very large
- Retroarticular process of articular absent
- Premaxillary teeth D- or U-shaped in cross-section with median ridge on caudal surface
- Premaxillary teeth smaller than lateral teeth
- Humerus greatly reduced; femur-humerus ratio between 2.8 and 3.5
- Fibular cranial tubercle distal to cranial expansion composed of two longitudinal ridges
- Dorsal surface of metatarsal III crescentic and restricted to caudal portion of the metatarsus dorsal surface
The following features are known in derived tyrannosaurids, are absent in potential outgroups (except some possible non-tyrannosaurid tyrannosauroids), but cannot be evaluated at present in the more fragmentary basal forms. Thus, these may represent synapomorphies of Tyrannosauridae, Tyrannosaurinae, or some subgroup thereof:
- Premaxillary tooth row arcade more mediolaterally than rostrocaudally oriented
- Lateral nuchal crest formed by parietals present, at least twice as tall as foramen magnum vertical height
- Pair of tab-like processes on supraoccipital wedge
- Basisphenoidal sphenoidal sinus deep, with large foramina
- Prominent muscular fossae on dorsal surface of palatine absent
- Rostral portion of fused vomers expanded to greater than twice shaft width
- Supradentary fused to coronoid, restricting lateral motion of intramandibular joint (Hurum and Currie 2000)
- Distal caudal neural spines axially elongate
- Acromial expansion well-developed, more than twice scapula midshaft width
- Scapula contributes more than half of the glenoid
- Scapular-humerus ratio between 2.2 and 2.5
- Metacarpal III very reduced and bears no digit
- Ilium long, slight shorter than femur
- Horizontal medial shelf from preacetabular blade to sacral ribs
- Broad ventral hook-like projection from preacetabular blade of ilium
- Pronounced midline crest on ilium
- Supracetabular crest on ilium reduced
- Pubic boot enlarged, approximately one-half pubis (or femur) length
- Ischium shaft long but more slender and shorter than pubis
Please see the page on Tyrannosaurid Systematics for characters supporting the subclades within the tyrant dinosaurs shown above.
Discussion of Phylogenetic Relationships
At the present time there are no published numerical phylogenetic analyses of tyrannosaurid relationships. The tree above is from work by Holtz in press (a, b). The synapomorphies supporting these relationship are discussed in detail on the Tyrannosaurid Systematics page.
Previous studies of tyrant dinosaur systematics have been concerned more with taxonomic issues of lumping and splitting rather than recovery of the phylogenetic relationships within the clade (e.g, Russell 1970, Maleev 1974, Paul 1988, Carpenter 1992). Russell (1970) presented one of the most extensive reviews of the Tyrannosauridae. His arrangement was as follows:
- Albertosaurus
- A. libratus
- A. sarcophagus
- A. lancensis
- Daspletosaurus torosus
- Tarbosaurs bataar
- Tyrannosaurus rex
This study was the first to recognize Daspletosaurus and was influential in including the late Campanian Age Gorgosaurus libratus within the genus Albertosaurus. This practice was followed by various other authors (e.g., Paul 1988, Carpenter 1992, Holtz 1994, 1995, Carr 1999), although the justification for the union of Albertosaurus sarcophagus and A. libratus was the shared absence of the derived features found in Daspletosaurus, Tarbosaurus, and Tyrannosaurus. Holtz (in press a, b) found that a few derived cranial characters do support union of Albertosaurus and Gorgosaurus outside of all other tyrannosaurines, but that equally parsimonious trees unite Gorgosaurus and the more robust tyrannosaurines to the exclusion of Albertosaurus: see Tyrannosaurid Systematics.
Gorgosaurus lancensis, known only from a single skull from the Hell Creek Formation of Montana, was also provisionally moved into Albertosaurus by Russell (1970), Paul (1988), and Carpenter (1992). Bakker et al. (1988) referred this specimen to its own genus, Nanotyrannus. Carr (1999), however, demonstrated that this individual is most likely a juvenile specimen of Tyrannosaurus rex.
Carpenter's (1992) arrangement was largely similar to that of Russell, but included Tarbosaurus bataar in Tyrannosaurus and introduced the new taxon Maleevosaurus novojilovi (previously Gorgosaurus novojilovi) for a smaller specimen from Mongolia. The latter is regarded a subadult specimen of Tarbosaurus bataar by Carr (1999) and Holtz (in press a, b).
Paul (1988) differed with Russell (1970) and Carpenter (1992) in expanding Tyrannosauridae to include the more plesiomorphic aublysodontines and (provisionally) Indosuchus raptorius, a form now considered a member of the ceratosaurian clade Abelisauridae. Paul included Shanshanosaurus houyanshanensis as a species of Aublysodon, Daspletosaurus as a subgenus of Tyrannosaurus, and provisionally included Alectrosaurus olseni in Albertosaurus. Additionally, Paul recognized a distinct species Albertosaurus arctunguis for specimens regarded by other authors (Russell 1970, Carpenter 1992, Holtz in press a) as belonging to A. sarcophagus, and a new species Albertosaurus megagracilis for a specimen considered by Carr (1999) and Holtz (in press a, b) to be a subadult Tyrannosaurus rex. Paul's arrangement is as follows:
- Aublysodontinae
- Aublysodon
- A. mirandus
- A. molnari
- A. houyanshanensis
- Aublysodon
- Tyrannosaurinae
- ?Indosuchus raptorius
- Alioramus remotus
- Albertosaurus
- ?A. olseni
- A. libratus
- A. sarcophagus
- A. arctunguis
- A. megagracilis
- ?A. lancensis
- Tyrannosaurus
- T. (Daspletosaurus) torosus
- T. (Tyrannosaurus)
- T. (T.) bataar
- T (T.) rex
Horner et al. (1992) briefly mention a new taxon from the upper Two Medicine Formation of Montana. Based on a the distribution of a few derived features, they proposed the following topology:
====== Daspletosaurus torosus ![[Extinct]](/tree/ToLimages/extinct.gif) =====|
| === Two Medicine tyrannosaurine
===|
=== Tyrannosaurus rex
=====|
| === Two Medicine tyrannosaurine
===|
=== Tyrannosaurus rex
However, those authors did not include the Asian taxon Tarbosaurus bataar in their (admittedly preliminary) study; Holtz (in press a, b) found that when that Mongolian species is included, the relationship of the unnamed Two Medicine form becomes unresolved with regards to Daspletosaurus and the Tarbosaurus-Tyrannosaurus clade.
Aublysodontinae is included within Tyrannosauridae by some authors (Paul 1988, Currie et al. 1990, Holtz 1994, in press a, b, Kirkland et al. 1997, Cifelli et al. 1997, 1999), while others use a more restricted Tyrannosauridae and use the term "Aublysodontidae" for Aublysodon and its allies (Molnar and Carpenter 1989, Carpenter 1992): see Taxonomy for the taxonomic definitions used here.
Taxonomy
For the purposes of discussion here, the following provisional taxonomic definitions will be used:
- Tyrannosauridae, all descendants of the most recent common ancestor of Tyrannosaurus and Aublysodon
- Tyrannosaurinae, Tyrannosaurus and all taxa closer to Tyrannosaurus than to Aublysodon
- Aublysodontinae, Aublysodon and all taxa closer to Aublysodon than to Tyrannosaurus
- Tyrannosauroidea, all taxa closer to Tyrannosaurus than to Ornithomimus or to Deinonychus
However, these definitions must be provisional, as the type species of Aublysodon (A. mirandus) is known only from isolated premaxillary teeth, while A. molnari may represent a different genus. Material currently referred to Alectrosaurus olseni is under study; this material may eventually provide a much more useful anchor taxon for Tyrannosauridae.
(Note: the taxonomic definitions provided by Sereno (1998) are problematic, as in that taxonomy Tyrannosauridae represents Tyrannosaurus and all taxa closer to it than to Alectrosaurus, Aublysodon, and Nanotyrannus. The latter "taxon", however, appears to be a juvenile specimen of T. rex (Carr 1999).)
See Holtz (in press a, b) for further discussion of these taxonomic definitions.
Stratigraphic and Geographic Distribution
Several partially known theropod taxa have been considered closer to tyrannosaurids than to all other known clades of dinosaur: see Tyrannosauroidea. Zinke (1998) described premaxillary teeth similar in form to those of tyrant dinosaurs from the Late Jurassic (Kimmeridgian Age) of Portugal. These specimens differ in form from those of known tyrannosaurids in being squatter mediolaterally compared to crown height. The Portuguese teeth are considerably older than other tyrannosaurid taxa, and might be from non-tyrannosaurid tyrannosauroids, basal ornithomimosaurs, or some other as-yet unknown taxon.
Manabe (1999) has described a premaxillary tooth from the Jobu Formation (Early Cretaceous, ?Valanginian-?Barremian Age) of Japan, identical in form to those of typical tyrannosaurids. Teeth similar in form to those of Alectrosaurus are reported from the latest Albian-earliest Cenomanian Mussentuchit Member of the Cedar Mountain Formation of Utah (Kirkland et al. 1997; Cifelli et al. 1997, 1999). These teeth are thus older than all skeletal material unquestionably assignable to Tyrannosauridae. The latter are known at present from the Late Cretaceous Epoch. Alectrosaurus olseni represents the oldest named tyrannosaurid: depending on the age of the Iren Dabasu and Bayn Shire Formations, it may be as old as the Cenomanian to as young as the early Campanian Age (Currie and Eberth 1993). The remaining tyrannosaurid taxa are of late Campanian or Maastrichtian Age. Other than the Portuguese teeth, tyrannosaurid fossils are restricted to central and eastern Asia and North America.
References
Bakker, R. T., Williams, M., and Currie, P. J. 1988. Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana. Hunteria 1(5): 1-30.
Barsbold, R. 1983. [Carnivorous dinosaurs from the Cretaceous of Mongolia]. Sovmestnaia Sovetsko-Mongol'skaia Palentologischeskaia Ekspeditsiia Trudy 19: 1-117. (In Russian with English summary)
Carpenter, K. 2000. Evidence of predatory behavior by carnivorous dinosaurs. Gaia 15: 135-144.
Carr, T. D. 1999. Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria). Journal of Vertebrate Paleontology 19: 497-520.
Carrano, M. T. 1998. What, if anything, is a cursor? Categories versus continua for determining locomotor habit in mammals and dinosaurs. Journal of Zoology (London) 247: 29-42.
Carroll, R. L. 1988. Vertebrate Paleontology and Evolution. W. H. Freeman, New York. 698 pp.
Chin, K., Tokaryk, T. T., Erickson, G. M., and Calk, L. C. 1998. A king-sized coprolite. Nature 393: 680-682.
Cifelli, R. L., Kirkland, J. I., Weil, A., Deino, A. L., and Kowallis, B. J. 1997. High-precision 40Ar/39Ar geochronology and the advent of North America's Late Cretaceous terrestrial fauna. Proc. Natl. Acad. Sci. USA 94: 11163-11167.
Cifelli, R. L., Nydam, R. L., Gardner, J. D., Weil, A., Eaton, J. G., Kirkland, J. I., and Madsen, S. K. 1999. Medial Cretaceous vertebrates from the Cedar Mountain Formation, Elliot County, Utah: the Mussentuchit Local Fauna. In: Gillette, D. D. (ed.) Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publications 99-1: 219-242.
Colbert, E. H. 1955. Evolution of the Vertebrates. Wiley, New York. 479 pp.
Collinvaux, P. 1978. Why Big Fierce Animals Are Rare: an Ecologist's Perspective. Princeton University Press, Princeton. 256 pp.
Cope, E. D. 1892. On the skull of the dinosaurian Laelaps incrassatus Cope. Proceedings of the American Philosophical Society 30: 241.
Currie, P. J. 2000. Possible evidence for gregarious behavior in tyrannosaurids. Gaia 15: 271-277.
Currie, P. J. and Eberth, D. A. 1993. Palaeontology, sedimentology and palaeoecology of the Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia, People's Republic of China. Cretaceous Research 14: 127-144.
Currie, P. J., Rigby, J. K., Jr., and Sloan, R. E. 1990. Theropod teeth from the Judith River Formation of southern Alberta, Canada. In: Currie, P. J., and Carpenter, K. (eds.) Dinosaur Systematics: Approaches and Perspectives. Cambridge University Press, Cambridge. Pp. 107-125.
Erickson, G. M. 1995. Split carinae on tyrannosaurid teeth and implications of their development. J. Vert. Paleontol. 15: 268-274.
Erickson, G. M., and Olson, K. H. 1996. Bite marks attributable to Tyrannosaurus rex: preliminary description and implications. Journal of Vertebrate Paleontology 16: 175-178.
Erickson, G. M., Van Kirk, S. D., Su, J., Levenston, M. E., Caler, W. E., and Carter, D. R. 1996. Bite-force estimation for Tyrannosaurus rex from tooth-marked bones. Nature 382: 706-708
Farlow, J. O. 1993. On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large carnivorous dinosaurs. American Journal of Science 293-A: 167-199.
Farlow, J. O., Brinkman, D. L., Abler, W. L., and Curie, P. J. 1991. Size, shape, and serration density of theropod dinosaur lateral teeth. Modern Geology 16: 161-198.
Farlow, J . O., Smith, M. B., and Robinson, J. M. 1995. Body mass, bone "strength indication," and cursorial potential of Tyrannosaurus rex. Journal of Vertebrate Paleontology 15: 713-725.
Feduccia, A. 1999. The Origin and Evolution of Birds. Second Edition. Yale University Press, New Haven.
Forster, C. A., Sampson, S. D., Chiappe, L. M., and Krause, D. W. 1998. The theropod ancestry of birds: new evidence from the Late Cretaceous of Madagascar. Science 279: 1915-1919.
Gauthier, J. A. 1986. Saurischian monophyly and the origin of birds. In: Padian, K. (ed.). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8: 1-55.
Halstead, L. B., and Halstead, J. 1981. Dinosaurs. Blandford Press, Poole, England. 170 pp.
Holtz, T. R., Jr. 1994. The phylogenetic position of the Tyrannosauridae: implication for theropod systematics. Journal of Paleontology 68: 1100-1117.
Holtz, T. R., Jr. 1995. The arctometatarsalian pes, an unusual structure of Cretaceous Theropoda (Dinosauria: Saurischia). Journal of Vertebrate Paleontology 14: 408-519.
Holtz, T. R., Jr. 1996. Phylogenetic taxonomy of the Coelurosauria (Dinosauria: Theropoda). Journal of Paleontology 70: 536-538.
Holtz, T. R., Jr. 2000. A new phylogeny of the carnivorous dinosaurs. Gaia 15: 5-62.
Holtz, T. R., Jr. in press a. Tyrannosauridae. In: Weishampel, D. B., Dodson, P., and Osmólska, H. (eds.). The Dinosauria. Second Edition. University of California Press, Berkeley.
Holtz, T. R., Jr. in press b. The phylogeny and taxonomy of the Tyrannosauridae. In: Carpenter, K., and Tanke, D. (eds.). Mesozoic Vertebrate Life. Indiana University Press.
Holtz, T. R., Jr. in press c. Evidence for the predatory nature of Tyrannosaurus rex and other tyrant dinosaurs. DinoFest III Proceedings.
Horner, J. R. 1994. Steak knives, beady eyes, and tiny little arms (a portrait of T. rex as a scavenger). In: Rosenberg, G. D., and Wolberg, D. L. (eds.). DinoFest Proceedings. Paleontological Society Special Publications 7: 157-164.
Horner, J. R., and Dobb, E. 1997. Dinosaur Lives: Unearthing an Evolutionary Saga. Harcourt Brace and Company, San Diego. 244 pp.
Horner, J. R., and Lessem, D. 1993. The Complete T. rex: How Stunning New Discoveries Are Changing Our Understanding of the World's Most Famous Dinosaur. Simon and Schuster, New York. 239 pp.
Huene, F. von. 1923. Carnivorous Saurischia in Europe since the Triassic. Bulletin of the Geological Society of America 34: 449-458.
Huene, F. von. 1926. The carnivorous Saurischia in the Jura and Cretaceous formations, principally in Europe. Revista Museo de La Plata 29: 35-167.
Huene, F. von. 1932. Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte. Monographie für Geologie und Palaeontologie (series 1) 4: 1-361.
Hurum, J. H., and Currie, P. J. 2000. The crushing bite of tyrannosaurids. Journal of Vertebrate Paleontology 20: 619-621.
Kirkland, J. I., Britt, B., Burge, D. L., Carpenter, K., Cifelli, R., DeCourten, F., Eaton, J., Hasiotis, S., and Lawton, T. 1997. Lower to Middle Cretaceous dinosaur faunas of the central Colorado Plateau: a key to understanding 35 million years of tectonics, sedimentology, evolution and biogeography. BYU Geology Studies 42: 69-103.
Lambe, L. B. 1904. On Dryptosaurus incrassatus (Cope) from the Edmonton Series of the North West Territory. Contributions to Canadian Palaeontology 3: 1-27.
Lambe, L. B. 1917. The Cretaceous theropodous dinosaur Gorgosaurus. Memoirs of the Geological Survey of Canada 100: 1-84.
Leidy, J. 1856. Notice of remains of extinct reptiles and fishes, discovered by Dr. F. V. Hayden in the badlands of the Judith River, Nebraska Territory. Proceedings of the Academy of Natural Sciences of Philadelphia 8: 72-73.
Makovicky, P. J., and Sues, H.-D. 1998. Anatomy and phylogenetic relationships of the theropod dinosaur Microvenator celer from the Lower Cretaceous of Montana. American Museum Novitates 3240:1-27.
Maleev, E. A. 1974. [Gigantic carnosaurs of the family Tyrannosauridae]. Sovmestnaia Sovetsko-Mongol'skaia Palentologischeskaia Ekspeditsiia Trudy 1: 132-191. (In Russian with English summary)
Manabe, M. 1999. The early evolution of the Tyrannosauridae in Asia. Journal of Paleontology 73: 1176-1178.
Marsh, O. C. 1890. Description of new dinosaurian reptiles. American Journal of Science (series 3) 39: 81-86.
Marsh, O. C. 1892. Notes on Mesozoic vertebrate fossils. American Journal of Science (series 3) 44: 171-176.
Matthew, W. D., and Brown, B. 1922. The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta. Bulletin of the American Museum of Natural History 46: 367-385.
Molnar, R. E. 1991. The cranial morphology of Tyrannosaurus rex. Palaeontograph. Abt. A 217: 137-176.
Molnar, R. E. 2000. Mechanical factors in the design of the skull of Tyrannosaurus rex. Gaia 15: 193-218.
Molnar, R. E., and Carpenter, K. 1989. The Jordan theropod (Maastrichtian, Montana, U.S.A.) referred to the genus Aublysodon. Geobios 22: 445-454.
Molnar, R. E., Kurzanov, S. M., and Dong Z.-M. 1990. Carnosauria. In: Weishampel, D. B., Dodson, P., and Osmólska, H. (eds.). The Dinosauria. University of California Press, Berkeley. Pp. 169-209.
Novas, F. E. 1992. La evolucion de los dinosaurios carnivoros. In: Sanz, J. L., and Buscalioni, A. D. (eds.). Los Dinosaurios y Su Entorno Biotico. Actas del Segundo Curso de Paleontologia en Cuenca. Instituto "Juan de Valdes", Cuenca. Pp. 125-163.
Osborn, H. F. 1905. Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bulletin of the American Museum of Natural History 21: 259-265.
Osborn, H. F. 1906. Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (second communication). Bulletin of the American Museum of Natural History 22: 281-296.
Osborn, H. F. 1912. Crania of Tyrannosaurus and Allosaurus. Memoirs of the American Museum of Natural History 1: 1-30.
Osborn, H. F. 1916. Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus. Bulletin of the American Museum of Natural History 35: 733-771.
Parks, W. A. 1933. New species of dinosaurs and turtles from the Belly River Formation of Alberta. University of Toronto Studies (Geological Series) 34: 1-33.
Paul, G. S. 1988. Predatory Dinosaurs of the World. Simon and Schuster, New York. 464 pp.
Paul, G. S. 2000. Limb design, function and running performance in ostrich-mimics and tyrannosaurs. Gaia 15: 257-270.
Pérez-Moreno, B. P., Sanz, J. L., Sudre, J., and Sigé, B. 1993. A theropod dinosaur from the Lower Cretaceous of southern France. Revue de Paléobiologie, Volume spécial 7: 173-188.
Pérez-Moreno, B. P., Sanz, J. L., Buscalioni, A. D., Moratalla, J. J., Ortega, F., and Rasskin-Gutman, D. 1994. A unique multitoothed ornithomimosaur dinosaur from the Lower Cretaceous of Spain. Nature 370:363-367.
Romer, A. S. 1945. Vertebrate Paleontology. 2nd Edition. Univ. Chicago Press, Chicago. 687 pp.
Russell, D. A. 1970. Tyrannosaurs from the Late Cretaceous of western Canada. National Museum National Science Publications in Palaeontology 1: 1-34.
Sereno, P. C. 1997. The origin and evolution of dinosaurs. Annual Review of Earth and Planetary Sciences 25: 435-489.
Sereno, P. C. 1998. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontolgie Abhandlungen 20: 41-83.
Sereno, P. C. 1999. The evolution of dinosaurs. Science 284: 2137-2147.
Sereno, P. C., Wilson, J. A., Larsson, H. C. E., Dutheil, D. B., and Sues, H.-D. 1994. Early Cretaceous dinosaurs from the Sahara. Science 266: 267-271.
Sereno, P. C., Dutheil, D. B., Iarochene, M., Larsson, H. C. E., Lyon, G. H., Magwene, P. M., Sidor, C. A., Varricchio, D. J., and Wilson, J. A. 1996. Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation. Science 272: 986-991.
Steel, R. 1970. Saurischia. Handbuch der Paläoherpetologie 14: 1-87.
Walker, A. D. 1964. Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs. Philosophical Transactions of the Royal Society of London Series B 248: 53-134.
Zinke, J. 1998. Small theropod teeth from the Upper Jurassic coal mine of Guimarota (Portugal). Palaontologische Zeitschrifte 72: 179-189.
Information on the Internet
- Tyrannosaurus rex Exposition. UCMP Berkeley.
- Tyrannosaurus Was Not A Fast Runner. John R. Hutchinson and Mariano Garcia.
- Sue at the Field Museum. Information about Sue, the largest, most complete, best preserved Tyrannosaurus rex found to date.
- Tyrannosaurus Was Not A Fast Runner. John R. Hutchinson and Mariano Garcia.
Title Illustrations

| Scientific Name | Tyrannosaurus rex, Triceratops horridus |
|---|---|
| Location | North America |
| Comments | The late Maastrichtian North American Tyrannosaurus rex guarding a carcass of the ceratopsid Triceratops horridus |
| Specimen Condition | Fossil -- Period: late Maastrichtian |
| Behavior | guarding a carcass |
| Copyright |
© 1998 Michael Skrepnick

|
About This Page
Thomas R. Holtz, Jr.
University of Maryland, College Park, Maryland, USA
Page copyright © 2000
All Rights Reserved.
- First online 29 August 2000
Citing this page:
Holtz, Jr., Thomas R. 2000. Tyrannosauridae. Tyrant dinosaurs. Version 29 August 2000 (under construction). http://tolweb.org/Tyrannosauridae/15896/2000.08.29 in The Tree of Life Web Project, http://tolweb.org/

