



Percomorpha
G. David Johnson and E. 0. Wiley




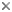
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Percomorphs (more than 14,450 species in about 250 families) are the crown group of the spiny-rayed fishes, and best represent what Nelson (1989) called the “bush at the top.” Percomorpha, sensu Johnson and Patterson (1993) comprise a group of orders forming the Smegmamorpha and five additional groups, traditionally treated as orders: Perciformes, Scorpaeniformes, Pleuronectiformes, Dactylopteriformes, and Tetraodontiformes.
The Smegmamorpha, with over 2,000 species in 37 families, are an unlikely and diverse assemblage comprising spiny and swamp eels, the Synbranchiformes (100 species in 3 families), grey mullets, the Mugiliformes (70 species in 1 family), pygmy sunfishes, the Elassomatiformes (6 species in 1 family), sticklebacks, pipefishes and allies, the Gasterosteiformes (275 species in 11 families), and the speciose silversides, flyingfishes, killifishes, and allies, the Atherinomorpha (1550 species in 21 familes and 3 orders). As recognized by Johnson and Patterson (1993), smegmamorphs are united by a single specialization, a unique attachment of the first epineural at the tip of a prominent transverse process on the first vertebra, but several additional specializations are shared by most of them. To date, smegmamorph monophyly has not been challenged by a comprehensive morphological analysis. Molecular analyses have failed to capture smegmamorph monophyly, although major components of the group have been recovered (e.g., Wiley et al. 2000; Chen et al., 2003; Miya et al., 2000, 2003, 2005).
The remaining percomorph groups, Johnson and Patterson's "perciforms and their immediate relatives" comprise over 12,550 species in approximately 210 families. They are currently classified in five separate orders, Perciformes (10,035, species in about 160 familes), Scorpaeniformes (scorpionfishes, sculpins, and allies; 1,470, species in 25 families), Dactylopteriformes (flying gurnards; 7 species in 1 family), Pleuronectiformes (flatfishes; 680, species in 14 families), and Tetraodontiformes (triggerfishes, puffers, and allies; 360, species in 9 families), however Johnson and Patterson (1993) saw no reason to exclude the latter four from the Perciformes and thought it likely that one or more are nested within that assemblage. Even without those four, the Perciformes are by far the largest and most diverse order of vertebrates, and although they include a number of freshwater groups, most species are marine, and they represent the dominant component of coral reef and inshore fish faunas.
Taxonomically, the Perciformes represent a catch-all assemblage of familes and suborders whose relationships have not been shown convincingly to lie elsewhere. Although there is good support for monophyly of about half the suborders, others remain poorly defined, most notably the largest, Percoidei (3200, species in about 80 families), another catch-all assemblage or "wastebasket" group, for which not a single diagnostic character has been proposed. Percoids are usually referred to as "perchlike fishes," and although this general physiognomy characterizes many familes (e.g., freshwater perches - Percidae, sunfishes - Centrarchidae, seabasses - Serranidae, and others), percoids encompass a wide variety of body forms, from the deep-bodied moonfishes - Menidae, butterflyfishes - Chaetodontidae, and more, to very elongate, eel-like forms such as bearded snakeblennies - Notograptidae, and bandfishes - Cepolidae. For lists and discussions of perciform suborders and percoid families, see Johnson (1993), Johnson and Gill (1998), and Nelson (2006), each of which, not surprisingly, differ somewhat in definition and composition of groups.
Characteristics
Synapomorphies
- Rod-like interarcual cartilage between widely separated uncinate processes of first epibranchial and second pharyngobranchial. (Johnson and Patterson, 1993; Travers, 1981). Absence of the interarcual cartilage in a number of percomorph groups was hypothesized to be secondary by Johnson and Patterson (1993) based on a wide separation of the tips of the uncinate processes of the first epibranchial and second pharyngobranchials, which are closely approximated in the primitive condition. Small, ovoid, non-homologous cartilages between the two uncinate processes are found in some myctophids, melamphaids, and beryciforms s. s.
- Second ural centrum absent (Johnson and Patterson, 1993). A separate second ural centrum is evident in adults of at least some members of all pre-percomorph lineages but lacking in adults of all members of the Percomorpha.
- Five or fewer hypurals (Johnson and Patterson, 1993). Apparently independently derived in melamphaids, berycids, holocentrids, diretmids, and zeiforms. Batrachoidiformes, Lophiiformes, Ophiidiformes also have five or fewer hypurals, but character conflict here is resolved by their inclusion in percomorphs.
- Pelvic fins with fewer than six soft rays (Johnson and Patterson, 1993). A variable character hypothesized to have been achieved independently within most lineages of basal acanthomorphs. Reversals to more rays seen in some smegmamorphs and pleuronectiforms. Pattern of loss may clarify the homology of lost rays in these groups; see Johnson and Patterson (1993) for discussion.
- Transforming ctenoid scales (Roberts, 1993; Johnson and Patterson, 1993). Several Smegamorph groups lack this character including atherinomorphs, gasterosteiforms, and synbranchiforms. Absent in tetradontiforms and sporadically in many perciforms (Roberts, 1993).
- Free pelvic radials lost (Johnson and Patterson, 1993).
- Point of origin of all but the first two epineurals displaced ventrally, and the distal parts of all epineurals displaced ventrally into the horizontal septum (Johnson and Patterson, 1993). A smilar pattern is found in Aphredoderus, gadiforms, one stephanoberyciform, some zeiforms, and the beryciform Monocentrus. Rejected as a holacanthopterygian character by Johnson and Patterson (1989) on the grounds of parsimony.
- Seventeen principal caudal fin rays in I,8,7,I pattern (Fujita, 1990). Reduction of rays common in percomorphs, increases are rare. Johnson and Patterson (1993) interpreted the 15 principal rays of Zeiformes as a convergence.
Discussion of Phylogenetic Relationships
As initially defined by Rosen (1973) Percomorpha was essentially the equivalent of Greenwood et al.’s (1966) Acanthopterygii minus Atherinomorpha. As such, it comprised beryciforms, perciforms, and the groups previously placed between and beyond those two, such as lampridiforms, zeiforms, gasterosteiforms, scorpaeniforms, pleuronectiforms and tetraodontiforms. Rosen presented no characters in support of his Percomorpha, nor have any valid ones been presented subsequently (possible exceptions proposed by Stiassny, 1990 and Roberts, 1993 were rejected by Johnson and Patterson, 1993).
The major goal of Johnson and Patterson’s (1993) analysis was to sort out basal lineages of acanthomorphs and revise the composition of Percomorpha to represent a monphyletic group diagnosed by derived characters. In the process, they erected a new, putatively monophyletic assemblage, Smegmamorpha, which, together with "the perciforms and their immediate relatives," the Scorpaeniformes, Pleuronectiformes, Dactylopteriformes, and Tetraodontiformes, constituted the newly defined Percomorpha. They identified eight evolutionary innovations of the Percomorpha, all of which are homoplasious at some level.
Although monophyly of Johnson and Patterson’s Percomorpha has not been challenged subsequently with morphological analyses, it is considered tenuous, particularly in view of our uncertainty of the composition and intrarelationships of the "perciforms and their immediate relatives," (which Johnson and Patterson did not test) and strong doubts about paracanthopterygian monophyly. As discussed on the Acanthopterygii page, to date, no molecular analyses have captured a monophyletic Percomorpha without the inclusion of two or three “paracanthopterygian” lineages, Batrachoidiformes, Lophiiformes and Ophidiiformes.
The Perciformes are almost certainly paraphyletic, and there is evidence that the other orders may be related to suborders within Perciformes. For example, with regard to the Scorpaeniformes, Mooi and Gill (1995) classified them within Perciformes based on a specific pattern of epaxial musculature shared with many perciforms. Mooi and Johnson (1997) suggested that the perciform champsodontids may be scorpaeniforms, and more recent morphological and molecular studies (e.g., Imamura and Yabe, 2002; Smith and Wheeler, 2004) support two distinct scorpaeniform lineages, Scorpaenoidei and Cottoidei, each with a different relationship to other percomorphs. Clearly the "percomorph problem" is the oustanding and most complex issue in our understanding of the evolutionary history of teleost fishes, and we may expect many changes in the classification of percomorphs with increased understanding of the relationships among these groups.
References
Chen, W.-J., C., Bonillo, and G. Lecointre. 2003. Repeatability of clades as a criterion of reliability: a case study for molecular phylogeny of Acanthomorpha (Teleostei) with larger number of taxa. Mol. Phylo. Evol. 26(2):262-288.
Fujita, K. 1990. The caudal skeleton of teleostean fishs. Tokai University Press. 897 pp.
Greenwood, P. H., D. E. Rosen, S. H. Weitzman, and G. S. Myers. 1966. Phyletic studies of teleostean fishes, with a provisional classification of living forms. Bull. Am. Mus. Nat. Hist. 131:339-456.
Imamura, I. and M. Yabe. 2002. Demise of the Scorpaeniformes (Actinopterygii: Percomorpha): an alternative phylogenetic hypothesis. Bull. Fish. Sci. Hokkaido Univ. 53(3):107-128.
Johnson, G. D. 1993. Percomorph phylogeny: progress and problems. Bull. Mar. Sci. 52(1):3-28.
Johnson, G. D. and A. C. Gill. 1998. Perches and their allies. Pp. 181-194 in Encyclopedia of Fishes. A comprehensive guide by international experts, 2nd ed. (J. R. Paxton and W. N. Eschmeyer, eds.). Academic Press, San Diego.
Johnson, G. D. and C. Patterson. 1993. Percomorph phylogeny: a survey of acanthomorphs and a new proposal. Bull. Mar. Sci. 52(1):554-626.
Miya, M. A, A. Kawaguchi, and M. Nishida. 2001. Mitogenomic exploration of higher teleostean phylogenies: a case study of moderate-scale evolutionary genomics with 38 newly determined complete mitochondrial DNA sequences. Mol. Biol. Evol. 18(11)1993-2009.
Miya, M., H. Takeshima, H. Endo, N. B. Ishiguro, J. G. Inous, T. Mukai, T. P. Satoh, M. Yamagucki, A. Kawaguchi, K. Mabuchi, S. M. Shirai, and M. Nishida. 2003. Major patterns of higher teleost phylogenies: a new perspective based on 100 complete mitochondrial DNA sequences. Mol. Phylo. Evol. 26:121-138.
Miya, M., Satoh, T.P., and Nishida. 2005. The phylogenetic position of toadfishes (Order Batrachoidiformes)in the higher ray-finned fishes as inferred from partitioned Bayesian analysis of 102 whole mitochondrial sequences. Biol. Jour. Linn. Soc. 85:289-306
Mooi, R. D., and A. C. Gill. 1995. Association of epaxial musculature with dorsal-fin pterygiophores in acanthomorph fishes, and its phylogenetic significance. Bull. Nat. Hist. Mus., London, Zool. Ser. 61:121-137.
Mooi, R. D. and G. D. Johnson. 1997. Dismantling the trachinoidei: evidence of a scorpaenoid relationship for the Champsodontidae. Ichthyol. Res. 44(2):143-176.
Nelson, G. 1989. Phylogeny of major fish groups. In: The Hierarchy of Life (B. Fernholm, K. Bremer, L. Brundin, H. Jornvall, L. Rutberg, and H.-E. Wanntorp, eds.). Elsevier, Amsterdam: 325-336.
Nelson, J. S. 2006. Fishes of the World, 4th Edition. Wiley-Interscience, New York.
Roberts, C. 1993. The comparative morphology of spined scales and their phylogenetic significance in the Teleostei. Bull. Mar. Sci. 52:60-113.
Rosen, D. E. 1973. Interrelationships of higher euteleostean fishes. In: Interrelationships of Fishes (P. H. Greenwood et al., eds.). J. Linnean Soc. (Zool.) 53, Suppl. 1:397-513.
Smith, W. L., and W. C. Wheeler. 2004. Polyphyly of the mail-cheeked fishes (Teleostei: Scorpaeniformes): evidence from mitochondrial and nuclear sequence data. Mol. Phylogenet. Evol.
Stiassny, M. L. J. 1990. Notes on the anatomy and relationships of the bedotiid fishes of Madagascar, with a taxonomic revision of the genus Rheocles (Atherinomorpha: Bedotiidae). Am. Mus. Nov. 2979:1-33.
Travers, R. A. 1981. The interarcual cartilage; a review of its development, distribution and value as an indicator of phylogenetic relationships in euteleostean fishes. J. Nat Hist. 15:853-871.
Wiley, E. O., G. D. Johnson, and W. W. Dimmick. 2000. The interrelationships of acanthomorph fishes: a total evidence approach using morphological and molecular data. Biochem. Syst. Evol. 28(2000):319-350.
Title Illustrations

| Scientific Name | Selene vomer |
|---|---|
| Location | Sarasota Aquarium, Florida, USA |
| Specimen Condition | Live Specimen |
| Source | silverfish from left |
| Source Collection | Flickr |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial-ShareAlike License - Version 2.0. This media file is licensed under the Creative Commons Attribution-NonCommercial-ShareAlike License - Version 2.0.
|
| Copyright | © 2005 Kai Schreiber |

| Scientific Name | Scorpaena papillosa |
|---|---|
| Location | Sydney, New South Wales, Australia |
| Specimen Condition | Live Specimen |
| Source | Moody Red Rock Cod |
| Source Collection | Flickr |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial-ShareAlike License - Version 2.0.
|
| Copyright | © 2006 Stuart Hamilton |
About This Page

Smithsonian Institution, Washington, D. C., USA
E. 0. Wiley
University of Kansas, Lawrence, Kansas, USA
Correspondence regarding this page should be directed to G. David Johnson at and E. 0. Wiley at
Page copyright © 2006 and E. 0. Wiley
All Rights Reserved.
- First online 09 January 2007
- Content changed 09 January 2007
Citing this page:
Johnson, G. David and E. 0. Wiley. 2007. Percomorpha. Version 09 January 2007 (under construction). http://tolweb.org/Percomorpha/52146/2007.01.09 in The Tree of Life Web Project, http://tolweb.org/

